| ADVERTISEMENT |
|
|
|
| Special Report: A Lecture by Nobel Laureate Dr. Paul Greengard |
||||||||||
|
Dr. Paul Greengard is an internationally-recognized researcher, currently investigating molecular signalling in the brain and its ramifications for neurological and psychiatric disorders. He has a long and distinguished teaching career – 22 years of teaching Pharmacology and Psychiatry at the Albert Einsten College of Medicine and the Yale University School of Medicine – as well as prestigious research positions as the director of Biochemistry at Geigy Research Laboratories (1959-67) and as Head of the Laboratory of Molecular and Cellular Neuroscience at Rockefeller University (1983 – present). Recently, he was honored with the 2000 Nobel Prize in the area of Physiology and Medicine, along with colleagues Arvid Carlsson and Eric Kandel, for discoveries concerning signal transduction in the nervous system. Recently, Dr. Greengard graciously agreed to lecture at Emory University, hosted by the institution’s Center for Behavioral Neuroscience. His hour-long talk covered some of his laboratories most recent investigations of a particular protein molecule (called DARPP-32) found in dopamine synapses, and what importance it holds for correct and incorrect dopamine signalling in the brain. As dopamine is one of the key neurotransmitters in the neuropathology of schizophrenia, Dr. Greengard’s research holds many future implications for greater understanding and treatment of the disorder. DARPP-32 appears to both regulate and be regulated by many neurotransmitters that also affect schizophrenia-like behavior; likewise, Greengard's lab also reports that DARPP-32 levels are reduced in the post-mortem brains of schizophrenia patients, as compared to normal controls. Although this does not prove that DARPP-32 is the cause of schizophrenia, it seems to play a role in the essential signaling pathways that affect the disease course, and as such could be a future therapeutic target. The full lecture is available in .mp3 format for downloading; however, readers without background in advanced neuromolecular physiology may find the abridged lecture summary below more helpful. Lecture by Dr. Paul Greengard at Emory University (April 25, 2005) – Highlights and Explanations
Cyclic-AMP: Also called cAMP, this is a protein molecule that is synthesized within neurons in response to second messenger signalling (after the binding of neurotransmitters to special types of receptors on the neuron membrane surface). Cyclic-AMP molecules activate other molecules known as protein kinases, which generally act to make the neuron more sensitive to neurotransmitter signalling (i.e. strengthening the synaptic connection) and/or turn on genes inside the neuron that synthesize more neurotransmitter receptors and make new synaptic connections. DARRP-32 ( dopamine and cyclic AMP-regulated phosphoprotein): A protein molecule that is found inside dopamine neurons. It is activated by the binding of dopamine to the neuron synapse, and also by the action of cyclic AMP molecules (which are synthesized within the neuron in response to the binding of several different neurotransmitters, including glutamate). DARRP-32 action is regulated by phosphorylation (the transfer of a phosphate molecule onto the protein itself) at certain locations, which influences the shape of the protein molecule and allows it to exert various actions within the neuron. NMDA Receptors: A special kind of glutamate receptor, which requires three things to activate – a molecule of glutamate binding to the active site, a molecule of glycine (or glycine-like compounds) binding to a regulatory site, and the depolarization of the synaptic membrane (depolarization occurs when a neuron is excited and ready to send a signal). NMDA receptors are present all over the brain, and are especially important for cellular learning and memory. Protein Kinases and Phosphatases: Enzymes in neurons that are activated by second-messenger neurotransmitter signalling. They work in opposition to each other – kinases phosphorylate (attach a phosphate molecule) onto molecular targets in a neuron, while phosphatases remove phosphate groups. One common protein kinase found in neurons, activated by the presence of lots of cyclic-AMP molecules, is called protein kinase A (abbreviated PKA). Slow transmission, or Second Messenger, Signalling: A response that occurs when neurotransmitters bind to a special kind of post-synaptic receptor on the membrane called a G-protein coupled receptor. A chain of related events take place within the neuron itself, generally leading to the activation of protein molecules and/or the activation of genes inside the neuron that make more receptors and synapses. One of the molecules that can be generated due to second messenger signalling is cyclic AMP. Ultimately, the effects of second messenger signals will make a certain neuron either more or less sensitive to neurotransmitter signals (telling it to fire or not to fire). These effects last hours to days, and sometimes longer. Synapses and Synaptic Signalling: See the following website - http://nobelprize.org/medicine/educational/synapse/intro.html - for an excellent introduction on how messages travel between neurons in the brain.
Note: the following material is summarized directly from Dr. Greengard’s own words, presented at Emory University on April 25, 2005.Direct quotes are identified by quotation marks; anything else is the author’s own words based on comments by Dr. Greengard. Because Dr. Greengard received the Nobel prize for his work in slow-transmission dopamine signalling in the brain, he begins with an overview of different types of signalling, and then goes into greater detail on his own work with dopamine slow-transmission signalling and the post-synaptic proteins involved in that process. Finally, he presents research data that give some clinical applications for his work in such areas as schizophrenia and substance abuse. Special thanks to Dr. Greengard and to the organizers at Emory University for permission to record and reproduce the words of the lecture.
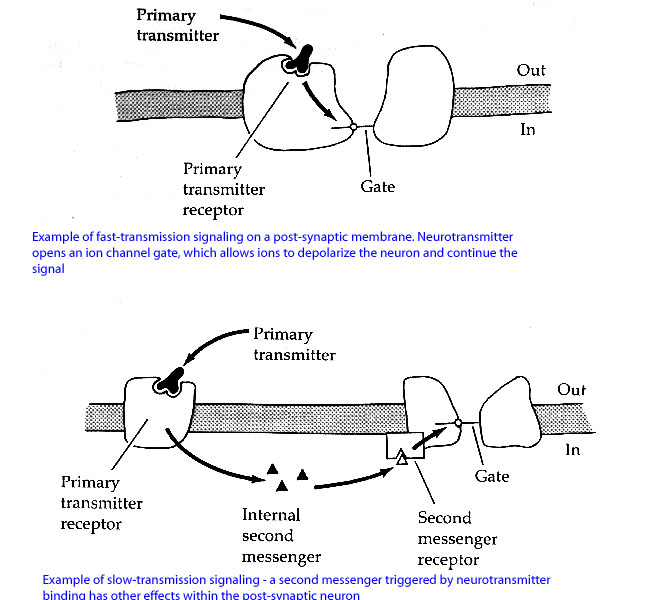
An Overview of Signalling in the Brain: There are two kinds of signalling in the brain – fast transmission signalling and slow transmission signalling. The type of signal that occurs depends not on the neurotransmitter so much as the type of receptor available for binding on the post-synaptic membrane (for example, glutamate can initiate both fast and slow transmission signals, because it can bind to different types of receptors). An example of fast transmission might be something like the following: a molecule of Neurotransmitter X binds to a receptor on the post-synaptic membrane, which opens an ion channel in the membrane that allows ions (i.e. sodium, potassium, chlorine) to flow in or out of the neuron. This ion flow leads to rapid neuron events – either depolarization (that neuron is likely to fire) or hyperpolarization (that neuron is much less likely to fire). The whole sequence of events in fast transmission, from neurotransmitter binding to ion traffic across the membrane, takes less than 1 millisecond. Slow transmission, on the other hand, involves a different type of receptor called a G-protein coupled receptor (see essential vocabulary for a detailed explanation of G-protein coupled receptors). Slow transmission signalling within the neuron takes several seconds, and ultimately results in second messengers (molecules inside the neuron) becoming active and phosphorylating target proteins and receptors. These cellular activities change the longer-term sensitivity of the neuron, usually making it more or less sensitive to fast-transmission signalling in the future. Below is a slightly modified image of how fast transmission (top) and slow transmission (bottom) signalling might take place. (Original web source: http://hebb.mit.edu/courses/8.515/lecture1/sld037.htm)
Of the hundreds of neurotransmitters that are involved with slow-transmission signalling, dopamine has been studied the most, and it is the first of the systems that Dr. Greengard’s research team studied. The four main brain disorders associated with dopamine transmission are: Parkinson’s disease, schizophrenia, ADHD, and Alzheimer’s. The main protein molecule studied by Greengard and his team is the Dopamine-and-cAMP-regulated-phosphoprotein (abbreviated: DARPP 32). Depending on which neurotransmitter is regulating it (and it is regulated by many different neurotransmitters), it may either inhibit cellular phosphatases, or inhibit cAMP dependent kinases. These two activities of DARPP have exactly opposite effects on the overall sensitivity of the neuron to neurotransmitter signals.
A large overview of a modulatory signalling pathway involving DARPP may look something like the following: glutamatergic cell bodies (cells which use glutamate as a neurotransmitter) project from the brain cortex to the neo-striatum, an area of the limbic system. Glutamate excites DARPP-containing neurons in the neo-striatum, which then send their signals to the substantia nigra (another part of the limbic system) onto noradrenaline and dopamine neurons. The dopamine neurons release dopamine back onto the original neo-striatal neurons (those that contain the DARPP molecule). This cyclical model allows a feed-back loop back to the dopaminergic neurons, which, through slow-transmission and DARPP protein involvement, can actually modify their signalling capacity over time. This type of model may be a mechanism through which slow-synaptic (dopamine) signalling regulates fast-transmission (glutamate) signalling. The next question is: how do slow-signalling dopamine receptor neurons regulate glutamate receptor neurons? First, it’s important to know that there are several different types of receptors involved. Dopamine has at least four receptors that we know about: in this situation, we are talking about D1 and D2 receptors. Glutamate has three different types of receptors, AMPA and NMDA receptors (fast-transmission receptors) and metabotropic (slow-transmission) receptors. Leaving glutamate out for the moment, let’s examine how DARPP can have post-synaptic effects that enhance a neuron’s sensitivity to dopamine transmission. Consider the following model, proposed by Dr. Greengard’s research: dopamine released from a dopaminergic neuron binds to the D1 receptor, a slow-transmission receptor. Dopamine binding activates the second messenger cAMP, which activates Protein Kinase A (PKA), which phosphorylates DARPP proteins at a certain amino acid on its structure (specifically, at Threonine residue 34). When phosphorylated at this position, DARPP inhibits a phosphatase protein (PP1). Inhibiting PP1 allows PKA to have a longer active life, and one of the other protein targets of PKA is dopamine receptors on the membrane surface Thus, the overall effects of the D1 DARRP pathway is to enhance the neuron’s sensitivity to dopamine signalling. In contrast, dopamine binding to a neuron with D2 receptors has an opposite effect: the second messenger phosphorylates DARPP in a different position, causing it to act as a protein kinase inhibitor. This decreases the overall phosphorylation within the neuron, and decreases the neuron’s receptivity to signalling. This model is the first example of how neurotransmitter transmission can regulate phosphorylation activity inside the post-synaptic neuron. Dr. Greengard’s research also shows that every compound known to influence the physiology of dopaminergic cells (for example: caffeine, cocaine, antipsychotics, antidepressents, alcohol, and many other drugs of abuse) also has an effect on DARPP. This was discovered mostly through observing the effects of these pharmacologically-active compounds on DARPP-32 knockout-mice (mice without the genes to synthesize DARPP-32 in their brains). Other neurotransmitters also seem to act at least partially through the actions of DARRP: for example, adenosine, serotonin, and GABA act on DARPP to increase intracellular neuron phosphorylation, while glutamate can act on DARPP through the AMPA-type receptor to decrease intracellular phosphorylation
Studies of DARPP-knockout mice give at least one example. In normal mice, high levels of circulating dopamine (or a dopamine agonist, such as cocaine) have distinct behavior and locomoter effects. Greengard et al. have shown that in DARPP-32 knockout mice, the effects of administered dopamine were greatly reduced or abolished altogether. This has extreme implications for schizophrenia research, given that the effects of overactive dopamine neurons are often used as a model for at least the positive symptom-spectrum of schizophrenia disorders. An important point is that DARPP works throuhg an amplification mechanism. The sequence of steps – neurotransmitter to second messenger to DARPP phosphorylation to DARPP effects – all involve molecules that have the capacity to activate not just one target, but many. Therefore, at each step in the chain there can be an exponential increase in the number of active molecules. This means that any modification to even just one molecule (in this case, DARPP) could have significant behavioral effects. Such behavioral effects controlled by DARPP have also been demonstrated by Greengard et al. For example, haloperidol and clozapine (used to treat schizophrenia symptoms) appear to both work through a similar DARPP-regulated mechanism – that is, increasing phosphorylation of DARPP by affecting the upstream pathway with receptor signaling. In the case of haldol, the molecule blocks the D2 receptor pathway, which increases DARPP phosphorylation. (Note: although haldol and clozapine both increase phosphorylation activity of PKA and DARPP-32, they have opposite effects on the activity of transcription factors within neurons. Therefore, even two pathways that converege at DARPP-32 may have divergent effects downstream). Prozac also increases DARPP phosphorylation. A phosphorylated DARPP 32 molecule is a potent inhibitor of a primary phosphatase in the brain (a phosphatase is a molecule that de-phosphorylates things). Therefore, a DARPP-32 molecule that is activated by phosphorylation (responding to a chemically induced signal cascade) actually helps that neuron signal longer and more efficiently. This is the suggested mechanism for how compounds like haloperidol, prozac, and clozapine exert their effects on dopamine neurons. Greengard presents further evidence that such therapeutic compounds work through DARPP modification. His lab found that neither DARPP-32 knockout mice, nor genetically altered mice with mutated DARPP-32 molecules had any reaction to Prozac administration. Greengard et al. suggested a more specific mechanistic model for how Prozac might work via DARPP in the brain. In areas such as the hippocampus, cortex, and striatum, Prozac helps prevent the reuptake of serotonin, allowing it to signal longer to the post-synaptic neuron. One of the normal actions of serotonin is to raise cyclic AMP levels within the post-synaptic neuron (via a receptor signaling cascade); increased cAMP levels activate PKA, which in turn activates DARPP-32 in a phosphatase-inhibiting role. Serotonin also activates a different signal cascade pathway that further decreases dephosphorylation of intracellular molecules via another mechanism. Activating these two mechanisms to decrease de-phosphorylation activity means that PKA can remain active longer, and exert intracellular effects such as increasing receptor sensitivity, activating transcription factors, and even eventually forming new synapses. Greengard’s lab has also tested the effects of hallucinogenic substances (LSD, PCP, etc) in mice unable to make a functional DARPP-32 molecule. In normal mice, hallucinogen administration causes specific behavioral effects, such as increased prepulse startle (a mouse that has been trained to pair a sound with a closely-followed electric shock will react to the sound even before the shock is administered – this is known as prepulse startle). In mice with non-functional DARPP-32 molecules, the behavioral effects of hallucinogens are similarly abolished. DARPP is clearly an important molecule to study for future schizophrenia therapy, since LSD and PCP induce some of the same symptoms in normal subjects as those seen in people with schizophrenia. What does this mean for understanding what might cause schizophrenia? Because DARPP-32 affects, and is affected by, so many of the neurotransmitters that also influence schizophrenia symptoms, an abnormal level or function of this protein in the brain might cause schizophrenia behavior. In a Clinical and Research News report (Psychiatric Times, Sept 2002), Greengard states: "DARPP-32 is a key regulatory protein, involved in controlling receptors, ion channels, and other physiological factors, and is activated and deactivated ultimately by neurotransmitters that are implicated in the development of schizophrenia...[a] reduction of DARPP-32, required for functions in the brain, could contribute to the cognitive dysfunction seen in the disease." In the same report, Greengard also reported on his lab research showing that DARPP-32 levels were reduced in the pre-frontal cortex of 14 post-mortem patients with schizophrenia, as compared with levels in gender- and age- matched control brains. How can changing just one molecule in brain neurons turn off a host of complex behaviors? One suggestions is because DARPP has many different sites of phosphorylation, it is an excellent candidate molecule to receive the downstream effects of signaling from all over the brain. For example, glutamate receptor activity initiates a signal cascade that affects DARPP in a different way; specifically, it makes DARPP a kinase inhibitor rather than a phosphatase inhibitor. In this role, DARPP acts in an opposite way from the pathway activated through D1 signalling. In short, the many phosphorylation sites of DARPP make it sensitive to many gradations of neurotransmitter signaling, as well as intracellular conditions and the activity of other intracellular molecules. This may be how just one molecule is able to exert such complex behavioral effects. When asked by an audience member about the specificity of the DARPP protein to dopamine neurons, Greengard hypothesized that DARPP 32 may have evolved early on as a mechanism to enhance D1 pathway signaling, but over the course of time was adopted by other neurotransmitter systems. For example, glutamatergic signaling in the brain has a modulatory effect on DARPP as well. In a recently published study from January 2005, Greengard et al tested the effects of glutamate administration on DARPP-32 containing neurons in the neostriatum of rat brains. They conclude that: “ glutamate activates at least five different signaling cascades with different time dependencies, resulting in complex regulation of protein kinase and protein phosphatase activities.” This provides more evidence that DARPP-32 activity may play a role in schizophrenia-spectrum symptoms, as many people with schizophrenia or schizophrenia-like symptoms have been shown to have aberrant glutamate signaling in their brains.
Dr. Paul Greengard’s research shows that some very complex behaviors can be altered (at least in a mouse model), simply by making very small changes to a single protein contained in neurons. He closed his lecture by stating that this finding "gives one cause for hope.” DARPP-32 provides a new therapeutic target for medication compounds, and the many ways it can be modified (through several different phosphorylation sites) makes it feasible that a wide range of symptoms may be correctable by affecting just this one molecule. View papers authored by Dr. Paul Greengard on pubmed.com:
Papers specifically on DARPP-32
|
ADVERTISEMENT
|